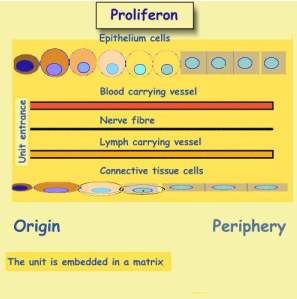
Epithelium cells are responsible for tasks
executed by the unit.. Connective tissue cells contribute the scaffold and
matrix. Nerve fibers provide communication with other units. Resources arecarried
by blood carrying vessels. Lymph vessels drain tissue fluid to
other units.
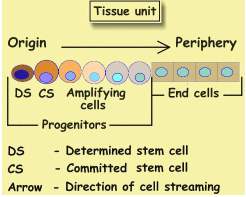
Epithelium cell kinetics
The tasks of each unit are carried out by epithelium, which supported by the
remaining components. There are two types of epithelia: Progenitors,
that divide, and non-dividing end-cells.. Progenitors like end-cells,
perform many metabolic functions, they differ from end-cells in their capability
to produce new cells. The cell at unit origin is called, Determined Stem
Cell (DS), Its more distant neighbor, is the Committed Stem Cell (CS).
|
|
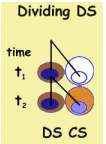 |
The scheme depicts an instantaneous picture of a unit in flux, directed
from origin to periphery. In order to appreciate it better let us follow a
DS at two time points: t1 and t2 , like
now and in one hour. Now DS is a single cell, and at t2 (next
hour) it divides into two cells. One offspring replaces the parent and remains
a DS, while the other becomes a CS which has to be placed beside the DS.
In order to enter its new location, all remote cells move outward and vacate
it. Thus, each progenitor cell division is accompanied by an outward steaming
of all remote cells. Cell streaming is linked with cell division, When progenitors
divide rapidly, remote cells travel faster, and vice versa. Cell flux and
turnover are linked.
All cells except the DS continually stream outward. Since during each DS division
one offspring replaces the parent cell, DS keeps its position at origin. Other
cells stream while dividing. Upon arriving at unit end, they die. Thus all cells
except DS, are called transitional cells and are short lived..
CS is the first transitional cell formed in the unit. As it streams it
ages, and its appearance changes. First turning into an amplifying
progenitor, "Amplifying" indicates that by repeated cell division
these cells increase the unit cell content. Further down along the cell
stream, a progenitor stops dividing , turning into an end-cell, and when
reaching unit periphery, it dies. This change of the cell is called differentiation.
In other words, as cells stream, they age and differentiate.
All cells in a tissue unit are descendants of one DS, belonging to one cell
clone. The tissue unit is therefore mono-clonal. Destroy the DS and the
entire unit gradually dies. It may live as long as transitional cells do. As
they approach periphery, the unit gradually dwindles away.
In the gut epithelium, a differentiating CS lives two days. In the
skin (epidermis) it lives three weeks. Cell streaming was described nearly in
all epithelia of the organism, e.g. gut,, skin liver, pancreas, salivary glands,
adrenal and kidney. The list of relevant publication is given below.We may safely
assume that wherever cells divide their turnover is structured in the same
way. All cells except heart cells, divide, and turn over. With the discovery
of dividing DS in the brain, even this organ turns over, and divided into tissue
units, that are structured in the same fashion as our simplified scheme.
|
 |
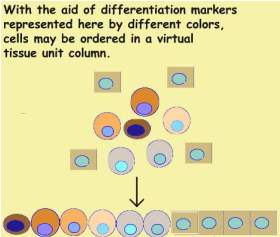
Not all units are so obvious like thos of the skin or gut. Bone marrow cells
are dispersed and it is difficult to imagine that they follow the above
scheme. Yet they do. Each cell carries a unique marker represented here by
its color. They may be sorted and arranged along a row exactly like
skin epithelium..Mathematically this observation may expressed as follows.
Since DS generates the entire unit. The various differentiation states
of cells may be defined by continuous transformations. All tissue units
are homeo-morphic.
Tissue unit behavior may now be summarized
by several rules:
1.
The unit is generated by its DS.
2. Cells
stream from their birth site to their graveyard.
3.
Velocity is proportional to cell division rate.
Embryogenesis
While a cell is the basic unit of the organism,
tissue unit is its basic building block. Our existence starts
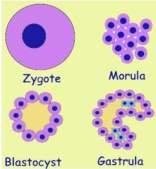
as a fertilized ovum, or zygote, our primordial stem cell. Its
nucleus contains a blueprint of the organism. A set of directives that will
serve for constructing it. Initially zygote is nourished by maternal fluids
of the womb. Resources enter the cell by diffusion. It rapidly divides
into a cell cluster, called morula. Since outer cells are first to
get their needs, less is left for inner cells. In order to get their needs
they have to push their way outward. The cluster turns into a balloon filled
with fluid, called blastocyst. Its skin is made of a single cell layer.
All cells get their needs and non is deprived anymore.
In the following developmental stage, called gastrula, some cells fold
into the balloon creating a small gut. Other cells form an inner layer. They
cannot rely anymore on getting resources by diffusion, and have to grow their
own vascular supply. This is achieved by creating tissue units with the five
ingredients mentioned above.
From now on embryo grows in a similar way as growing cities, by adding
quarters. Each tissue unit is like a city quarter. Its buildings are cells
and its infrastructure, vessels and nerve fibers,. Yet unlike real quarters,
the unit is oriented. A stem cell at its origin generates the entire unit.
Its progeny, the transitional cells stream to their graveyard.
Whenever a new unit is created, a stem cell buds off its parent. In
an adult that stopped growing, a dividing DS generates two different progeny,
a DS and a CS, which is called asymmetric stem cell division. In the
growing embryo, prior to forming a new unit the DS divides into two
DS , one maintains the old unit, while the other generates a new one. This
is known as symmetric stem cell division.
The
figure depicts a unit at four time points:
1. Parent
unit
2.
DS divides symmetrically creating a bud.
3.
Unit elongates to a 4-cell stage.
4. Unit
elongates to a 8-cell state.
Proliferon
Actually, unit budding is somewhat more
complicated. When an epithelial DS buds off its parent unit, it is joined
up by a connective tissue DS. Both get a supply of budding vessels, and nerve
fibers. As the unit elongates, its vessels and nerve fibers also elongate,
reminding of how new city quarters expand. This complex vas called Proliferon
.
|
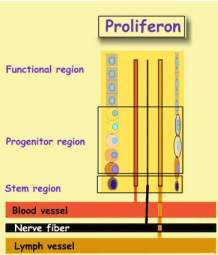 |
The scheme is an instantaneous image of a streaming proliferon. An
asymmetric epithelial DS division is accompanied by an asymmetric connective
tissue DS division. The two CS cells, start their voyage together, accompanied
by their infra-structure. Thus each DS division creates a proliferon
quantum that moves outward. In the urban context, formation of a CS is
like building a new house with its pipes and cables. Here, the house
gradually moves to the quarter periphery whereupon it is demolished. Not only
cells are being renewed, but vessels and nerve fibers as well.
In order to simplify the discussion, we shall concentrate on epithelial cells,
keeping in mind the proliferon.
Panta
Rhe (All streams)
Heraclitus once said: "All streams"
(panta rhe). And added:" you never step into the same river twice",
To which we may now add, you never meet the same individual twice, since all
his cells stream.
Tissue unit as JAVA program.
Zygote is the top most class in the organism. It is the root of the system.
DS cells are its subclasses. Transitional cells are objects
generated by DS. An asymmetric DS division instantiates an object.
When reaching tissue periphery, object dies and is cleared
by the garbage collector. DS class inherits its properties from the
zygote class. All DS cells in a given tissue belong to a package. A
symmetric DS division, is equivalent to the formation of a new class. Since
tissue packages differ from each other, their classes hide some variables,
and override some methods inherited from zygote. Household cell functions
are methods that cannot be overridden. Runtime environment emerges.
Neoplastic progression
Cancer is a systemic disease manifested by three features:
1. Cachexia, or body wasting.
2. Para-neoplasia, manifested by hormonal dysfunction and nerve conduction
disturbances.
3. Neoplasia, the tumor.
We shall restrict our discussion to tumors made of epithelial cells. When
treating laboratory animals with carcinogens, tumor evolution can be studied
from its very beginning. Most of the information described here was gathered
in mice and rats intestinal epithelium treated with carcinogens.
A tumor is initiated in a single tissue unit. A normal cell is transformed
into a neoplastic. Obviously this transformed cell has to be a DS, since
transformed transitional cells are soon washed out of the unit. Tumor
initiation is a stem cell event. Tumor, like other tissues, grows
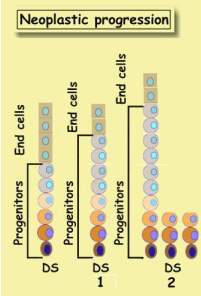
by addition of proliferons. The image depicts two stages of tumor formation
, starting with a healthy unit.
1 There are more progenitors and less end-cells.
2 The above
changes intensify. In addition, unit elongates, DS cells bud off, and create
new units.
Since progenitors become more abundant, more cells divide, their turnover rises,
and cell streaming accelerates. End-cell depletion, is known as Maturation
Arrest. The unit is less mature. With time these changes intensify, and
unit elongates. Gradually more and more new units appear.
These are the earliest changes in neoplastic (or tumor) evolution. They are
called also precursor lesions, exhibiting the salient features of a
tumor. Progenitor amplification. Particularly increased stem cell
abundance. In the gut it takes about five years, until a full
fledged neoplasm is detected.
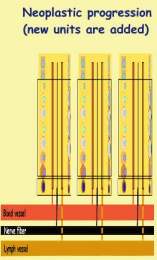
During evolution, tumor-proliferon changes its appearance. It grows in size,
multiplies its units, and becomes more and more immature. Tasks that were
hitherto performed by mature epithelia, are canceled (maturation arrest),
and replaced with new ones, that are now performed by progenitors. It has
to be stressed that progenitors like end-cells, perform many metabolic functions.
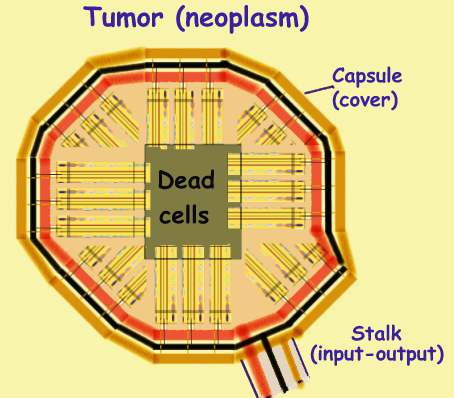
Tumor
Tumor is an organ with a new purpose, that evolves like an embryo by
proliferon addition. Generally DS cells are close to vessels from
which the budding proliferon draws its infra-structure. Due to its polarity,
the unit points away from adjacent vessels. Its origin rests near a
vessel and its periphery points away from it. As more units are added they
form a lump covered with a capsule through which vessels pass. Units whose
origins are a placed below the capsule point to the tumor center. Since
transitional cell stream inward where they die. Dead tumor cells accumulate
in the center.
As disease advances, DS cells enter adjacent vessels and travel to other tissues,
where they bud into proliferons called metastases.
Chemotherapy
A tumor grows in the same way as an embryo.
Its transitional cells are short lived, continually streaming to their graveyards.
Chemotherapy generally inhibits cell formation As transitional cells
continue dying, in the absence of new cells, tumor shrinks. DS
are spared, since resisting the poison. When chemotherapy is stopped, they
replenish their proliferons with transitional cells, and tumor grows
again. With repeated chemotherapy treatments, DS number rises so that
the entire tumor is made of DS (anaplasia), and resists chemotherapy.
Chemotherapy generally fails since enriching
resistant DS (clonogenic cells). Neoplastic DS resist chemotherapy in the same
way as normal DS do. One might raise the amount of poison and eliminate neoplastic
DS, yet this would destroy also all other DS and kill the patient. Since chemotherapy
fails to eliminate the tumor, it should be given solely for alleviating tumor
induced damage. Imagine a tumor that obstructs airways, and his size has
to be reduced. Treatment will be helpful if tumor is made mainly from transitional
cells. In order to minimize the threat of chemotherapy-resistance, poison should
be given sparingly. The smallest dose necessary for reducing tumor size, and
not more.
Tumor
is a sophisticated organ with a purpose whose nature medicine still fails
to understand.
Bibliography
1 Zajicek,G.;Yagil,Ch.;Michaeli,Y
The streaming submandibular gland.
The Anat. Rec. 213:150-158,1985.
2 Zajicek,G.;Oren,R.;Weinreb,Jr.
The streaming liver.
Liver 5:293-300,1985.
3 Zajicek,G.
The application of kinematic equations for the
study of cell turnover.
J. Theoret. Biol. 120:141-149,1986.
4 Zajicek G.,Ariel I.,Arber N.
The streaming adrenal cortex: direct evidence of
centripetal migration of
adrenocytes by estimation of cell turnover rate.
J. Endocr. 111: 447-482,1986.
5 Zajicek G. The time dimension in histology.
Methods of Inform. Med. 26:1-2,1987.
6 Arber N., Zajicek G., Ariel I.
The streaming liver II: Hepatocyte life history.
Liver 8:80-87, 1988.
7 Zajicek G., Ariel I., Arber N.
The streaming liver III: Littoral cells accompany the streaming hepatocyte.
Liver 8:213-218,1988.
8 Ariel I.,Kerem E.,Schwartz-Arad D.,Bartfeld E., Ron N.,Pizov G., Zajicek
G.
Nesidiodysplasia - A histologic entity ?
Human Pathol. 19:1215-1218,1988.
9 Schwartz-Arad D., Zajicek G., Bartfeld E.
Streaming Liver IV: DNA content of the hepatocyte increases with its
age.
Liver 9:93-99,1989.
10 Zajicek G.,Schwartz-Arad D.,Bartfeld E.
Streaming Liver V: Time and age dependent changes of hepatocyteDNA content,
following partial hepatectomy.
Liver 9:164-171 1989.
11 Breuer R,Zajicek G,Christensen TG, Lucey EC Snider, GL.
Cell kinetics of normal adult hamster bronchial epithelium in steady state.
Am. J. Respir. Cell Mol. Biol. 2:51-58,1990
12 Arber N, Zajicek G,
Streaming liver VI: Streaming intra-hepatic bile ducts.7
Liver 10:205-208,1990
13 Zajicek G, Schwartz-Arad D.
Streaming Liver VII: DNA Turnover in acinus zone 3.
Liver 10:137-140,1990.
14 Zajicek G.
Hepatocytes and intra-hepatic bile duct epithelium originate form a common
stem cell (letter to the editor).
Gastroenterol. 100:582, 1991.
15 Zajicek G, Arber N. Schwartz-Arad D, Ariel I.
Streaming Pancreas : Islet cell kinetics.
Diabet. Research. 13:121-125, 1990.
16 Zajicek G, Arber N, Schwatz-Arad D.
Streaming Liver VIII: Cell production rates following partial hepatectomy.
Liver 11:347-351,1991
17 Zajicek G
Time dimension in histopathology
Path. Res. Pract. 188:410-412,1992
18 Sipcic SR, Deutsch D, Zajicek G
Simulation of cancer progression in the colon on
a massively parallel processor (CM-2)
Proc. Fifth SIAM Conference on Parallel Processing for Scientific Computing.
Dongarra J,Kennedy K, Messina P, ßorensen DC, Voigt RG. Eds pp, 345-350,1991
19 Sipcic SR, Zajicek G
Kinetic analysis of epithelial cell migration in the colonon a massively
parallel processor (CM-2)
Proc. Sixth SIAM Conference on Parallel Processing for Scientific Computing
Sincovec R... [et al.] Eds Vol. 1 pp, 300-303, 1993
20 Sipcic SR, Zajicek G
Kinetic analysis of epithelial cell migration in the colon on a massively
parallel processor (CM-2) Comput. Biomed. Res. 26, 393-412,1993
21Zajicek G.
Streaming Organism: The Tissue Automat.
in: Computing with Biological Metaphors. Ed. R. Paton.Chapman & Hall
London, 1994.
|
Physician
|
 |
Philosopher
|
 |
|
Click on your icon
to follow your trail
|
Contents
Home